Model situacije "predator-plijen". Ravnoteža predator-plijen Matematički model konzervativnog plijena predatora
Kolmogorovljev model donosi jednu značajnu pretpostavku: budući da se pretpostavlja da to znači da u populaciji plijena postoje mehanizmi koji reguliraju njihovu brojnost čak i u odsutnosti predatora.
Nažalost, takva formulacija modela ne omogućuje odgovor na pitanje oko čega U zadnje vrijeme Postoji mnogo rasprava, koje smo već spomenuli na početku poglavlja: kako populacija predatora može izvršiti regulatorni utjecaj na populaciju plijena tako da cijeli sustav bude održiv? Stoga ćemo se vratiti na model (2.1), u kojem mehanizmi samoregulacije (na primjer, regulacija putem intraspecifične konkurencije) nedostaju u populaciji plijena (kao iu populaciji predatora); stoga je jedini mehanizam za regulaciju broja vrsta uključenih u zajednicu trofički odnos između grabežljivaca i plijena.
Ovdje (dakle, za razliku od prethodnog modela, naravno, rješenja (2.1) ovise o specifičnoj vrsti trofičke funkcije koja je pak određena prirodom grabežljivosti, tj. trofičkom strategijom grabežljivca i obrambena strategijažrtve. Sljedeća svojstva zajednička su svim ovim funkcijama (vidi sliku I):
Sustav (2.1) ima jednu netrivijalnu stacionarnu točku čije su koordinate određene iz jednadžbi
![]()
pod prirodnim ograničenjima.
Postoji još jedna stacionarna točka (0, 0), koja odgovara trivijalnoj ravnoteži. Lako je pokazati da je ta točka sedlo, a separatrise koordinatne osi.
Karakteristična jednadžba za točku ima oblik
![]()
Očito, za klasični model Volterra.
Stoga se vrijednost f može smatrati mjerom odstupanja razmatranog modela od Volterrinog modela.
![]()
stacionarna točka je fokus, a u sustavu se pojavljuju oscilacije; kada je suprotna nejednakost zadovoljena, postoji čvor i nema oscilacija u sustavu. Stabilnost ovog ravnotežnog stanja određena je uvjetom
tj. Značajno ovisi o vrsti trofičke funkcije predatora.
Uvjet (5.5) može se protumačiti na sljedeći način: za stabilnost netrivijalne ravnoteže sustava predator-plijen (a time i za postojanje tog sustava), dovoljno je da u blizini tog stanja relativni udio konzumiranog plijena od strane predatora povećava se s povećanjem broja plijena. Doista, udio plijena (od njihovog ukupnog broja) koji je pojeo grabežljivac opisuje se diferencijabilnom funkcijom, uvjet za koji se povećava (pozitivna derivacija) izgleda kao
![]()
Posljednji uvjet uzet u točki nije ništa drugo nego uvjet (5.5) za stabilnost ravnoteže. Uz kontinuitet, mora se ispuniti iu određenom susjedstvu točke. Dakle, ako je broj žrtava u tom susjedstvu, tada
Neka sada trofička funkcija V ima oblik prikazan na sl. 11, a (karakteristika beskralješnjaka). Može se pokazati da za sve konačne vrijednosti (budući da je konveksan prema gore)
odnosno za bilo koju vrijednost stacionarnog broja žrtava nejednakost (5.5) nije zadovoljena.
To znači da u sustavu s ovom vrstom trofičke funkcije ne postoji stabilna netrivijalna ravnoteža. Moguće je nekoliko ishoda: ili se broj plijena i grabežljivaca neograničeno povećava ili (kada putanja prolazi blizu jedne od koordinatnih osi) zbog slučajni razlozi broj plijena ili broj predatora postat će nula. Ako plijen ugine, nakon nekog vremena uginut će i grabežljivac, no ako grabežljivac prvi ugine, tada će se broj plijena početi eksponencijalno povećavati. Treća opcija - nastanak stabilnog graničnog ciklusa - je nemoguća, što se lako dokazuje.
Zapravo, izraz

u pozitivnom kvadrantu je uvijek pozitivan, osim ako ima oblik prikazan na sl. 11, a. Tada, prema Dulacovom kriteriju, u ovom području nema zatvorenih putanja i ne može postojati stabilan granični ciklus.
Dakle, možemo zaključiti: ako trofička funkcija ima oblik prikazan na Sl. 11, te tada predator ne može biti regulator koji osigurava stabilnost populacije plijena, a time i stabilnost cijelog sustava u cjelini. Sustav može biti stabilan samo ako populacija plijena ima vlastite interne regulacijske mehanizme, na primjer, intraspecifičnu konkurenciju ili epizootije. O ovoj mogućnosti regulacije već je bilo riječi u §§ 3, 4.
Prethodno je navedeno da je ova vrsta trofičke funkcije karakteristična za predatore insekata, čije su "žrtve" također obično insekti. S druge strane, zapažanja o dinamici mnogih prirodne zajednice tipovi "predator-plijen", u koje spadaju i vrste kukaca, pokazuju da ih karakteriziraju oscilacije vrlo velike amplitude i vrlo specifičnog tipa.
Obično nakon više ili manje postupnog porasta broja (koji se može odvijati monotono ili u obliku oscilacija s rastućom amplitudom) dolazi do oštrog pada (slika 14), a zatim se slika ponavlja. Očigledno se ovakva priroda dinamike brojnosti vrsta kukaca može objasniti nestabilnošću ovog sustava pri maloj i srednjoj brojnosti i djelovanjem snažnih intrapopulacijskih regulatora brojnosti pri velikoj brojnosti.

Riža. 14. Dinamika populacije australske ljupke Cardiaspina albitextura koja se hrani stablima eukaliptusa. (Iz članka: Clark L. R. Dinamika populacije Cardiaspina albitextura.-Austr. J. Zool., 1964., 12, br. 3, str. 362-380.)
Ako sustav “predator-plijen” uključuje vrste sposobne za dovoljno izazovno ponašanje(na primjer, grabežljivci su sposobni učiti ili žrtve mogu pronaći sklonište), tada je u takvom sustavu moguće postojanje stabilne netrivijalne ravnoteže. Ova izjava se dokazuje vrlo jednostavno.
Zapravo, trofička funkcija bi tada trebala imati oblik prikazan na Sl. 11, c. Točka na ovom grafu je tangentna točka ravne linije povučene iz ishodišta grafa trofičke funkcije. Očito, u ovoj točki funkcija ima maksimum. Također je lako pokazati da je uvjet (5.5) zadovoljen za sve. Posljedično, netrivijalna ravnoteža u kojoj je broj žrtava manji bit će asimptotski stabilna
Međutim, ne možemo ništa reći o tome koliko je veliko područje stabilnosti te ravnoteže. Na primjer, ako postoji nestabilan granični ciklus, tada ovo područje mora ležati unutar ciklusa. Ili druga opcija: netrivijalna ravnoteža (5.2) je nestabilna, ali postoji stabilan granični ciklus; u ovom slučaju možemo govoriti i o stabilnosti sustava predator-plijen. Budući da izraz (5.7) pri odabiru trofičke funkcije kao što je sl. 11, in može promijeniti predznak pri promjeni na , tada Dulacov kriterij ovdje ne radi i pitanje postojanja graničnih ciklusa ostaje otvoreno.

PA88 sustav, koji istovremeno predviđa vjerojatnost više od 100 farmakoloških učinaka i mehanizama djelovanja tvari na temelju njezine strukturne formule. Učinkovitost ovog pristupa planiranju probira je oko 800%, a točnost računalne prognoze je 300% veća od predviđanja stručnjaka.
Dakle, jedan od konstruktivnih alata za dobivanje novih znanja i rješenja u medicini je metoda matematičkog modeliranja. Proces matematizacije medicine česta je manifestacija prožimanja znanstvenih spoznaja, povećanja učinkovitosti liječenja i preventivnog rada.
4. Matematički model “predator-plijen”
Po prvi put u biologiji talijanski matematičar V. Volterra i njegovi kolege predložili su matematički model periodičnih promjena broja antagonističkih životinjskih vrsta. Model koji je predložio Volterra bio je razvoj ideje koju je 1924. godine zacrtao A. Lottka u knjizi “Elementi fizičke biologije”. Stoga je ovaj klasični matematički model poznat kao "Lottky-Volterra" model.
Iako su u prirodi odnosi antagonističkih vrsta složeniji nego u modelu, oni su ipak dobar nastavni model na kojemu se proučavaju osnovne ideje matematičkog modeliranja.
Dakle, problem: u nekom ekološki zatvorenom području žive dvije vrste životinja (na primjer, ris i zec). Zečevi (plijen) hrane se biljnom hranom, koja je uvijek dostupna u dovoljnoj količini (ovaj model ne uzima u obzir ograničene resurse biljna hrana). Risovi (grabežljivci) mogu jesti samo zečeve. Potrebno je utvrditi kako će se broj plijena i grabežljivaca mijenjati tijekom vremena u takvom ekološkom sustavu. Ako se populacija plijena povećava, povećava se i vjerojatnost susreta predatora i plijena, a sukladno tome, nakon određenog vremenskog odmaka, populacija predatora raste. Ovaj je dovoljan jednostavan model sasvim adekvatno opisuje interakciju između stvarnih populacija predatora i plijena u prirodi.
Sada počnimo sastavljanje diferencijalnih jednadžbi. Oko
označimo broj plijena s N, a broj predatora s M. Brojevi N i M su funkcije vremena t. U našem modelu uzimamo u obzir sljedeće faktore:
a) prirodna reprodukcija žrtava; b) prirodna smrt žrtava;
c) uništavanje žrtava jedenjem grabežljivaca; d) prirodno izumiranje predatora;
e) povećanje broja predatora zbog razmnožavanja uz prisutnost hrane.
Jer govorimo oŠto se tiče matematičkog modela, zadatak je dobiti jednadžbe koje bi uključivale sve predviđene faktore i koje bi opisivale dinamiku, odnosno promjenu broja predatora i plijena kroz vrijeme.
Neka se broj plijena i grabežljivaca promijeni za ∆N i ∆M tijekom nekog vremena t. Promjena broja žrtava ∆N tijekom vremena ∆t određena je, prvo, porastom kao rezultatom prirodne reprodukcije (koja je proporcionalna raspoloživom broju žrtava):
gdje je B koeficijent proporcionalnosti koji karakterizira stopu prirodnog izumiranja žrtava.
Izvođenje jednadžbe koja opisuje smanjenje broja plijena zbog njihove konzumacije od strane predatora temelji se na ideji da što se češće susreću, to se broj plijena brže smanjuje. Također je jasno da je učestalost susreta predatora i plijena proporcionalna i broju žrtava i broju predatora, dakle
Dijeleći lijevu i desna strana jednadžbu (4) na ∆t i prelazeći na granicu na ∆t→0, dobivamo diferencijalna jednadžba prva narudžba:
Kako biste riješili ovu jednadžbu, morate znati kako se broj predatora (M) mijenja tijekom vremena. Promjena broja grabežljivaca (∆M) određena je porastom zbog prirodnog razmnožavanja uz dovoljno hrane (M 1 = Q∙N∙M∙∆t) i smanjenjem zbog prirodnog izumiranja grabežljivaca ( M 2 = - P∙M∙∆ t):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Iz jednadžbe (6) možemo dobiti diferencijalnu jednadžbu:
Diferencijalne jednadžbe (5) i (7) predstavljaju matematički model "predator-plijen". Dovoljno je odrediti vrijednosti koeficijenta

za rješavanje problema mogu se koristiti elementi A, B, C, Q, P i matematički model.
Provjera i podešavanje matematičkog modela. U ovom laboratoriju
Uz izračun najpotpunijeg matematičkog modela (jednadžbe 5 i 7), predlaže se proučavanje jednostavnijih u kojima se nešto ne uzima u obzir.
Razmotrivši pet razina složenosti matematičkog modela, možete "osjetiti" fazu provjere i prilagodbe modela.
1. razina – u modelu se za “plijen” uzima u obzir samo njihova prirodna reprodukcija, nema “predatora”;
Razina 2 – model uzima u obzir prirodno izumiranje “plijena”, nema “predatora”;
Razina 3 – model uzima u obzir prirodnu reprodukciju “žrtava”
I izumiranje, nema “predatora”;
4. razina – model uzima u obzir prirodnu reprodukciju “žrtava”
I izumiranje, kao i pojedenost od strane “predatora”, ali broj “predatora” ostaje nepromijenjen;
Razina 5 – model uzima u obzir sve razmatrane čimbenike.
Dakle, imamo sljedeći sustav diferencijalnih jednadžbi:
gdje je M broj "predatora"; N – broj “žrtava”;
t – trenutno vrijeme;
A – stopa reprodukcije “žrtava”; C – učestalost susreta predatora i plijena; B – stopa izumiranja “žrtava”;
Q – razmnožavanje “predatora”;
P – izumiranje “predatora”.
1. razina: M = 0, B = 0; 2. razina: M = 0, A = 0; 3. razina: M = 0; 4. razina: Q = 0, P = 0;
Razina 5: potpuni sustav jednadžbi.
Zamjenom vrijednosti koeficijenata u svaku razinu, dobit ćemo različita rješenja, na primjer:
Za 3. razinu vrijednost koeficijenta M=0, dakle

rješavajući jednadžbu koju dobivamo
Isto tako za razine 1 i 2. Što se tiče 4. i 5. razine, ovdje je potrebno riješiti sustav jednadžbi metodom Runge-Kutta. Kao rezultat, dobivamo rješenje matematičkih modela ovih razina.
II. RAD STUDENATA TIJEKOM PRAKTIČNE NASTAVE
Vježba 1 . Kontrola usmenog govora i korekcija svladavanja teorijskog gradiva lekcije. Polaganje prijema u nastavu.
Zadatak 2. Izvođenje laboratorijskih radova, diskusija o dobivenim rezultatima, pisanje bilješki.
Završetak rada
1. S radne površine računala pozovite program "Laboratorija br. 6" dvostrukim klikom lijevom tipkom miša na odgovarajući prečac.
2. Dvaput kliknite lijevom tipkom miša na prečac "PREDATOR".
3. Odaberite prečac "PRED" i ponovite poziv programa lijevom tipkom miša (dvoklikom).
4. Nakon naslovnog zaslona pritisnite "ENTER".
5. Manekenstvo počinje s 1. razina.
6. Unesite godinu od koje će se model analizirati: na primjer, 2000
7. Odaberite vremenske intervale, na primjer, unutar 40 godina, nakon 1 godine (zatim nakon 4 godine).
2. razina: B = 0,05; N0 = 200;
3. razina: A = 0,02; B = 0,05; N = 200;
4. razina: A = 0,01; B = 0,002; C = 0,01; N0 = 200; M = 40; 5. razina: A = 1; B = 0,5; C = 0,02; Q = 0,002; P = 0,3; N0 = 200;
9. Pripremiti pisano izvješće o radu koje treba sadržavati jednadžbe, grafikone, rezultate proračuna karakteristika modela, zaključke o obavljenom radu.
Zadatak 3. Praćenje završne razine znanja:
a) usmeni izvještaj o obavljenom poslu laboratorijski rad; b) rješavanje situacijskih problema; c) računalno testiranje.
Zadatak 4. Zadatak za sljedeću lekciju: dio i tema lekcije, koordinacija tema za sažetke (opseg izvješća 2-3 stranice, vremensko ograničenje 5-7 minuta).
Prilagodbe koje je razvio plijen kako bi se suprotstavio predatorima pridonose razvoju mehanizama pomoću kojih predatori mogu prevladati te prilagodbe. Dugotrajni suživot predatora i plijena dovodi do stvaranja sustava interakcije u kojem su obje skupine stabilno očuvane na području istraživanja. Kršenje takvog sustava često dovodi do negativnih posljedica za okoliš.
Negativan utjecaj poremećaja koevolucijskih odnosa opaža se tijekom uvođenja vrsta. Konkretno, koze i zečevi, uneseni u Australiju, nemaju učinkovite mehanizme za regulaciju svog broja na ovom kontinentu, što dovodi do uništavanja prirodnih ekosustava.
Matematički model
Recimo da na nekom području žive dvije vrste životinja: zečevi (koji se hrane biljkama) i lisice (koje se hrane zečevima). Neka broj zečeva , broj lisica . Koristeći Malthusov model s potrebnim dopunama uzimajući u obzir jedenje zečeva od strane lisica, dolazimo do sljedeći sustav, koji nosi naziv Volterrinog modela - Poslužavnici:
\begin(cases) \dot x=(\alpha -c y)x;\\
\dot y=(-\beta+d x) y. \kraj(slučajevi)
Model ponašanja
Skupni stil života grabežljivaca i njihovog plijena radikalno mijenja ponašanje modela i daje mu veću stabilnost.
Obrazloženje: Skupni život smanjuje učestalost slučajnih susreta grabežljivaca i potencijalnog plijena, što potvrđuju i promatranja populacijske dinamike lavova i gnuova u parku Serengeti.
Priča
Model suživota dvoje biološke vrste(populacije) tipa “predator-plijen” naziva se i Volterra-Lotka model.
vidi također
Napišite recenziju članka "Sustav predator-plijen"
Bilješke
Književnost
- V. Volterra, Matematička teorija borba za egzistenciju. Po. s francuskog O. N. Bondarenko. Pod uredništvom i pogovorom Yu. M. Svirezheva. M.: Nauka, 1976. 287 str. ISBN 5-93972-312-8
- A. D. Bazykin, Matematička biofizika interakcijskih populacija. M.: Nauka, 1985. 181 str.
- A. D. Bazykin, Yu. A. Kuznetsov, A. I. Khibnik, Portreti bifurkacija (Bifurkacijski dijagrami dinamičkih sustava na ravnini) / Serija “Novo u životu, znanosti, tehnologiji. Matematika, kibernetika" - M.: Znanie, 1989. 48 str.
- P. V. Turchin,
Linkovi
Izvadak koji karakterizira sustav "predator-plijen".
“Šarmantno, šarmantno, [Ljupko, šarmantno”, rekao je princ Vasilij.“C"est la route de Varsovie peut être, [Ovo je Varšavska cesta, možda.] - rekao je princ Hippolyte glasno i neočekivano. Svi su se osvrnuli na njega, ne shvaćajući što je time želio reći. Princ Hippolyte također se osvrnuo s veselim iznenađenjem oko sebe.On kao i drugi nije shvaćao što znače riječi koje je izgovorio.Za vrijeme njegovog diplomatsku karijeru Ne jednom sam primijetio da su tako iznenada izgovorene riječi ispale vrlo duhovite, a za svaki slučaj izgovorio je te riječi, prve koje su mu pale na jezik. “Možda će uspjeti vrlo dobro”, pomislio je, “a ako ne uspije, moći će to tamo srediti.” Doista, dok je vladala neugodna tišina, ušlo je ono nedovoljno patriotsko lice, kome je Ana Pavlovna čekala da se obrati, a ona je, smiješeći se i mašući prstom Hipolitu, pozvala kneza Vasilija k stolu i dajući mu dvije svijeće i rukopis, zamolio ga da počne . Sve je utihnulo.
- Premilostivi care! - strogo izjavi knez Vasilij i obazre se okupljenima, kao da pita ima li tko što reći protiv ovoga. Ali nitko ništa nije rekao. “Moskovska matična stolica, Novi Jeruzalem, prima svoga Krista,” iznenada je naglasio svoje riječi, “kao majka u naručje svojih revnih sinova, i kroz nadolazeću tamu, gledajući sjajnu slavu tvoje moći, pjeva u oduševljenju : “Hosana, blagoslovljen onaj koji dolazi.” ! – Princ Vasilij izgovori ove posljednje riječi plačnim glasom.
Bilibin je pažljivo pregledao svoje nokte, a mnogi su, očito, bili plašljivi, kao da pitaju što im je krivo? Ana Pavlovna je šapatom ponavljala naprijed, kao starica koja moli za pričest: "Neka drski i drski Golijat..." šapnula je.
Knez Vasilij nastavi:
- “Neka drski i bahati Golijat s granica Francuske odnese do rubova Rusije smrtonosne strahote; krotka vjera, ta praćka ruskoga Davida, iznenada će se obrušiti na glavu njegovu krvoločnom ponosu. Ova slika svetog Sergija, drevnog revnitelja za dobro naše domovine, donosimo vam carskom veličanstvu. Bolestan sam jer me slaba snaga sprječava da uživam u vašem najljubaznijem razmišljanju. Šaljem tople molitve nebu, da Svemogući uzveliča trku pravednih i ispuni dobre želje Vašeg Veličanstva.”
– Quelle force! Quel stil! [Kakva moć! Kakav slog!] - čule su se pohvale čitatelju i književniku. Nadahnuti ovim govorom, gosti Ane Pavlovne dugo su razgovarali o situaciji u domovini i iznosili razne pretpostavke o ishodu bitke koja se trebala voditi neki dan.
"Vous verrez, [Vidjet ćete.]", reče Anna Pavlovna, "da ćemo sutra, na suverenov rođendan, dobiti vijesti." Imam dobar osjećaj.
Predosjećaj Ane Pavlovne doista se ostvario. Sljedećeg dana, tijekom molitve u palači povodom rođendana suverena, princ Volkonski je pozvan iz crkve i primio je omotnicu od princa Kutuzova. Ovo je bio izvještaj Kutuzova, napisan na dan bitke iz Tatarinove. Kutuzov je napisao da se Rusi nisu povukli ni koraka, da su Francuzi izgubili mnogo više od nas, da se užurbano javlja s bojnog polja, a da još nije uspio prikupiti najnovije podatke. Stoga je to bila pobjeda. I odmah, bez napuštanja hrama, stvorena je zahvalnost Stvoritelju za njegovu pomoć i pobjedu.
Predosjećaj Ane Pavlovne bio je opravdan i cijelo je jutro u gradu vladalo radosno praznično raspoloženje. Svi su pobjedu priznali kao potpunu, a neki su već govorili o hvatanju samog Napoleona, njegovom svrgavanju i izboru novog poglavara Francuske.
Daleko od posla i među uvjetima dvorskog života, događaji se vrlo teško odražavaju u svoj svojoj punini i snazi. Nehotice se opći događaji grupiraju oko jednog posebnog slučaja. Dakle, sada je glavna radost dvorjana bila koliko u činjenici da smo pobijedili, toliko iu činjenici da je vijest o ovoj pobjedi pala upravo na suverenov rođendan. Bilo je to kao uspješno iznenađenje. Vijesti Kutuzova također su govorile o ruskim gubicima, a među njima su bili Tučkov, Bagration i Kutajsov. Također, tužna strana događaja nehotice se u ovdašnjem petrogradskom svijetu grupirala oko jednog događaja – smrti Kutaisova. Svi su ga znali, suveren ga je volio, bio je mlad i zanimljiv. Na današnji dan svi su se susreli uz riječi:
- Kako se nevjerojatno dogodilo. Na samoj molitvi. A kakav gubitak za Kutais! Oh kakva šteta!
– Što sam ti rekao o Kutuzovu? - princ Vasilij je sada govorio s ponosom proroka. Uvijek sam govorio da je samo on sposoban poraziti Napoleona.
Ali sutradan nije bilo nikakvih vijesti iz vojske, a opći je glas postao alarmantan. Dvorjani su patili zbog patnje nepoznatog u kojem je vladar bio.
- Kakav je to položaj suverena! - govorili su dvorjani i više ga nisu veličali kao prethodnog dana, nego sada osuđivali Kutuzova, bivši uzrok vladareve brige. Na današnji dan princ Vasilij se više nije hvalio svojim štićenikom Kutuzovom, ali je šutio kada je u pitanju bio vrhovni zapovjednik. Osim toga, do večeri ovog dana, činilo se da se sve poklopilo kako bi stanovnike Sankt Peterburga bacilo u uzbunu i brigu: dodana je još jedna strašna vijest. Grofica Elena Bezukhova iznenada je umrla od toga strašna bolest, koje je bilo tako ugodno izgovoriti. Službeno, u velikim društvima, svi su govorili da je grofica Bezukhova umrla od strašnog napadaja angine pectorale [upala grla], ali u intimnim krugovima ispričali su detalje o tome kako je le medecin intime de la Reine d "Espagne [liječnik španjolske kraljice] propisao Heleni male doze nekakvog lijeka da bi proizveo određeni učinak; ali kako Helen, mučena činjenicom da je stari grof sumnjao na nju i činjenicom da joj muž kojem je pisala (taj nesretni razvratnik Pierre) nije odgovorio , iznenada popila ogromnu dozu lijeka koji joj je bio propisan i umrla u mukama prije nego što su joj mogli pružiti pomoć.Pričali su da su princ Vasilij i stari grof preuzeli Talijana, ali je Talijan pokazao takve bilješke nesretne pokojnice da je odmah bio pušten na slobodu.
RAČUNALNI MODEL “PREDATOR-ŽRTVA”
Kazačkov Igor Aleksejevič 1, Guseva Elena Nikolajevna 2
1 država Magnitogorsk Tehničko sveučilište ih. G.I. Nosova, Institut za graditeljstvo, arhitekturu i umjetnost, student 5. godine
2 Magnitogorsk Državno tehničko sveučilište nazvano po. G.I. Nosova, Institut za energetiku i automatizirane sustave, kandidat pedagoških znanosti, izvanredni profesor Odsjeka za poslovnu informatiku i informacijske tehnologije
anotacija
Ovaj članak posvećen je pregledu računalnog modela "predator-plijen". Provedeno istraživanje sugerira da modeliranje okoliša ima veliku ulogu u istraživanju okoliša. Ovo pitanje je višestruko.
RAČUNALNI MODEL "PREDATOR-ŽRTVA"
Kazačkov Igor Aleksejevič 1, Guseva Elena Nikolajevna 2
1 Državno tehničko sveučilište Nosov Magnitogorsk, Institut za građevinarstvo, arhitekturu i umjetnost, student 5. tečaja
2 Državno tehničko sveučilište Nosov Magnitogorsk, Institut za energetiku i automatizirane sustave, doktor pedagoških znanosti, izvanredni profesor na Odsjeku za poslovno računalstvo i informacijske tehnologije
Sažetak
Ovaj članak daje pregled računalnog modela "predator-žrtva". Studija sugerira da simulacija okoliša igra veliku ulogu u proučavanju okoliša. Ovaj problem je višestruk.
Ekološko modeliranje koristi se za proučavanje našeg okoliša. Matematički modeli se koriste u slučajevima kada nema prirodno okruženje i br prirodni objekti, pomaže u predviđanju utjecaja razni faktori na predmet koji se proučava. Ova metoda preuzima funkcije provjere, konstruiranja i interpretacije dobivenih rezultata. Na temelju takvih oblika modeliranje okoliša bavi se procjenom promjena u okolišu koji nas okružuje.
Trenutno se takvi oblici koriste za proučavanje okoliša oko nas, a kada je potrebno proučiti bilo koje od njegovih područja, koristi se matematičko modeliranje. Ovaj model omogućuje predviđanje utjecaja određenih čimbenika na predmet proučavanja. Svojedobno su tip "predator-plijen" predložili znanstvenici kao što su: T. Malthus (Malthus 1798, Malthus 1905), Verhulst (Verhulst 1838), Pearl (Pearl 1927, 1930), kao i A. Lotka ( Lotka 1925, 1927 ) i V. Volterra (Volterra 1926.) Ovi modeli reproduciraju periodički oscilatorni režim koji nastaje kao rezultat interspecifičnih interakcija u prirodi.
Jedna od glavnih metoda spoznaje je modeliranje. Osim što može predvidjeti promjene koje se događaju u okoliš, također pomaže u pronalaženju optimalnog načina rješavanja problema. Matematički modeli već se dugo koriste u ekologiji kako bi se utvrdili obrasci i trendovi u razvoju populacija, te pomogli u isticanju suštine opažanja. Izgled može poslužiti kao uzorak ponašanje, objekt.
Prilikom rekreacije objekata u matematičkoj biologiji koriste se predviđanja različitih sustava, predviđene su posebne individualnosti biosustava: unutarnja struktura pojedinca, uvjeti održavanja života, postojanost ekološki sustavi, zahvaljujući kojima se čuva vitalna aktivnost sustava.
Pojava računalnog modeliranja značajno je pomaknula granice istraživačkih mogućnosti. Pojavila se mogućnost multilateralne primjene teških oblika koji ne dopuštaju analitičko proučavanje; najnoviji smjerovi, kao i simulacijsko modeliranje.
Razmotrimo što je objekt modeliranja. “Objekt je zatvoreno stanište u kojem dolazi do interakcije dviju bioloških populacija: predatora i plijena. Dolazi do procesa rasta, izumiranja i razmnožavanja neposredno na površini staništa. Plijen se hrani resursima koji su prisutni u okolišu, dok se predatori hrane plijenom. U ovom slučaju, prehrambeni izvori mogu biti obnovljivi ili neobnovljivi.
Godine 1931. Vito Volterra je izveo sljedeće zakone odnosa grabežljivac-plijen.
Zakon periodičnog ciklusa - proces uništavanja plijena od strane grabežljivca često dovodi do periodičnih fluktuacija u veličini populacije obiju vrsta, ovisno samo o brzini rasta mesojeda i biljojeda, te o početnom omjeru njihovog broja.
Zakon očuvanja prosjeka - Prosječna brojnost svake vrste je konstantna, bez obzira na početnu razinu, pod uvjetom da su specifične stope povećanja populacije, kao i učinkovitost grabežljivosti, konstantne.
Zakon kršenja prosječnih vrijednosti - kada se obje vrste smanjuju proporcionalno broju, prosječna veličina populacije plijena se povećava, a grabežljivaca smanjuje.
Model grabežljivac-plijen poseban je odnos između grabežljivca i njegovog plijena, od kojeg oboje profitiraju. Preživljavaju najzdravije i najprilagođenije jedinke okolišnim uvjetima, tj. sve se to događa zahvaljujući prirodni odabir. U okruženju u kojem nema mogućnosti razmnožavanja, predator će prije ili kasnije uništiti populaciju plijena, uslijed čega će i ona sama izumrijeti.”
Na zemlji postoji mnogo živih organizama koji pod povoljnim uvjetima povećavaju broj srodnika na ogroman razmjer. Ta se sposobnost naziva: biotički potencijal vrste, tj. povećanje broja vrste u određenom vremenskom razdoblju. Svaka vrsta ima svoj biotski potencijal, na primjer velike vrste organizmi u godini mogu se povećati samo 1,1 puta, zauzvrat, organizmi manjih vrsta, kao što su rakovi itd. može povećati njihovu pojavu do 1030 puta, ali bakterije su i dalje više. U svakom od ovih slučajeva, stanovništvo će eksponencijalno rasti.
Eksponencijalni rast brojeva naziva se geometrijska progresija rast populacije. Ta se sposobnost može promatrati u laboratoriju kod bakterija i kvasaca. U izvanlaboratorijskim uvjetima eksponencijalni rast može se vidjeti na primjeru skakavaca ili kod drugih vrsta insekata. Takav porast broja vrsta može se primijetiti na onim mjestima gdje praktički nema neprijatelja, a hrane ima više nego dovoljno. Na kraju je došlo do porasta vrsta, nakon što se broj povećao na kratko vrijeme, rast populacije je počeo opadati.
Razmotrimo računalni model reprodukcije sisavaca koristeći Lotka-Volterra model kao primjer. Neka Na određenom području žive dvije vrste životinja: jeleni i vukovi. Matematički model promjene populacije u modelu Trays-Volterra:
Početni broj žrtava je xn, broj predatora je yn.
Parametri modela:
P1 – vjerojatnost susreta s predatorom,
P2 – koeficijent rasta predatora na račun plijena,
d – stopa smrtnosti predatora,
a – koeficijent porasta broja žrtava.
U zadatku obuke postavljene su sljedeće vrijednosti: broj jelena 500, broj vukova 10, stopa rasta jelena 0,02, stopa rasta vukova 0,1, vjerojatnost susreta s predatorom 0,0026, stopa rasta predatora na račun plijena bila je 0 ,000056. Podaci su izračunati za 203 godine.
Istražujemo utjecaj koeficijent porasta žrtava za razvoj dviju populacija, ostali parametri ostat će nepromijenjeni. U shemi 1 uočava se povećanje broja plijena, a zatim, s određenim kašnjenjem, uočava se povećanje broja grabežljivaca. Tada grabežljivci izbacuju žrtve, broj žrtava naglo opada, a nakon toga se smanjuje i broj grabežljivaca (slika 1).

Slika 1. Veličina populacije s niskom stopom nataliteta među žrtvama
Analizirajmo promjenu modela povećanjem nataliteta žrtve a=0,06. Na dijagramu 2 vidimo ciklički oscilatorni proces koji dovodi do povećanja broja obiju populacija tijekom vremena (slika 2).

Slika 2. Broj stanovnika prema prosječnom natalitetu žrtava
Razmotrimo kako će se dinamika populacije promijeniti kada visoka vrijednost stopa nataliteta žrtve je a=1,13. Na sl. 3 dolazi do naglog porasta broja obiju populacija, nakon čega slijedi izumiranje i plijena i grabežljivaca. To se događa zbog činjenice da se populacija plijena povećala do te mjere da su resursi počeli nestajati, što je rezultiralo izumiranjem plijena. Istrebljenje grabežljivaca događa se zbog činjenice da se broj plijena smanjio, a grabežljivcima je ponestalo resursa za preživljavanje.
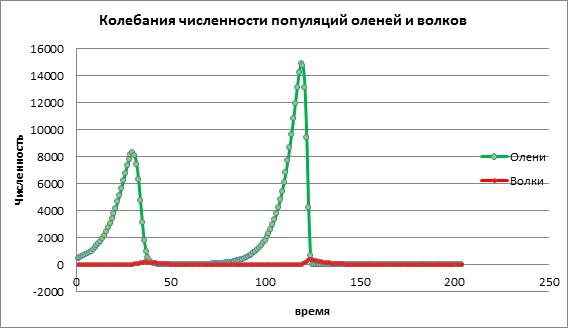
Slika 3. Veličina populacije s visokim stopama nataliteta među žrtvama
Na temelju analize podataka o računalnim eksperimentima, možemo zaključiti da nam računalno modeliranje omogućuje predviđanje veličine populacije i proučavanje utjecaja razni faktori o dinamici stanovništva. U gornjem primjeru ispitivali smo model predator-plijen, utjecaj nataliteta plijena na brojnost jelena i vukova. Mali porast populacije plijena dovodi do malog povećanja plijena, koji nakon određenog vremena grabežljivci uništavaju. Umjereno povećanje populacije plijena dovodi do povećanja veličine obje populacije. Visoki porast populacije plijena prvo dovodi do brzog porasta populacije plijena, to utječe na povećanje rasta grabežljivaca, ali zatim umnoženi grabežljivci brzo uništavaju populaciju jelena. Kao rezultat toga, obje vrste izumiru.
Modeli interakcije dva tipa
Volterrine hipoteze. Analogije s kemijskom kinetikom. Volterra modeli interakcija. Klasifikacija vrsta interakcija Natjecanje. Predator-plijen. Generalizirani modeli interakcija vrsta . Kolmogorov model. MacArthurov model interakcije dviju vrsta kukaca. Parametarski te fazni portreti Bazykinovog sustava.
Utemeljiteljem moderne matematičke teorije populacija s pravom se smatra talijanski matematičar Vito Volterra, koji je razvio matematičku teoriju bioloških zajednica, čiji su aparat diferencijalne i integro-diferencijalne jednadžbe.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Pariz, 1931). U narednim se desetljećima populacijska dinamika razvijala uglavnom u skladu s idejama iznesenima u ovoj knjizi. Ruski prijevod Volterrine knjige objavljen je 1976. pod naslovom: “Matematička teorija borbe za egzistenciju” s pogovorom Yu.M. Svirežev, koji ispituje povijest razvoja matematičke ekologije u razdoblju 1931.–1976.
Volterrina knjiga je napisana onako kako se pišu knjige o matematici. Prvo formulira neke pretpostavke o matematičkim objektima koji bi se trebali proučavati, a zatim provodi matematičko istraživanje svojstava tih objekata.
Sustavi koje je proučavao Volterra sastoje se od dva ili više tipova. U nekim slučajevima dolazi u obzir opskrba korištenom hranom. Jednadžbe koje opisuju interakciju ovih vrsta temelje se na sljedećim konceptima.
Volterrine hipoteze
1. Hrana je ili dostupna u neograničenim količinama, ili je njena nabava vremenski strogo regulirana.
2. Jedinke svake vrste umiru na takav način da konstantan udio postojećih jedinki umire u jedinici vremena.
3. Predatorske vrste jedu žrtve, a po jedinici vremena broj pojedenih žrtava uvijek je proporcionalan vjerojatnosti susreta jedinki ove dvije vrste, tj. umnožak broja predatora i broja plijena.
4. Ako postoji hrana u ograničenim količinama i nekoliko vrsta koje su je u stanju konzumirati, tada je udio hrane koju vrsta konzumira u jedinici vremena proporcionalan broju jedinki te vrste, uzet s određenim koeficijentom ovisno o vrste (modeli međuvrsne konkurencije).
5. Ako se neka vrsta hrani hranom koja je dostupna u neograničenim količinama, povećanje broja vrste u jedinici vremena proporcionalno je broju vrste.
6. Ako se vrsta hrani hranom koja je dostupna u ograničenim količinama, tada je njezino razmnožavanje regulirano brzinom potrošnje hrane, tj. po jedinici vremena, porast je proporcionalan količini pojedene hrane.
Analogije s kemijskom kinetikom
Ove hipoteze imaju bliske paralele s kemijskom kinetikom. U jednadžbama populacijske dinamike, kao iu jednadžbama kemijske kinetike, koristi se "princip sudara", kada je brzina reakcije proporcionalna umnošku koncentracija reagirajućih komponenata.
Doista, prema Volterrinim hipotezama, brzina postupak Izumiranje svake vrste proporcionalno je broju vrste. U kemijskoj kinetici to odgovara monomolekularnoj reakciji razgradnje određene tvari, au matematičkom modelu odgovara negativnim linearnim članovima na desnim stranama jednadžbi.
Prema pojmovima kemijske kinetike, brzina bimolekularne reakcije međudjelovanja između dviju tvari proporcionalna je vjerojatnosti sudara tih tvari, tj. proizvod njihove koncentracije. Isto tako, prema Volterrinim hipotezama, stopa razmnožavanja predatora (smrt plijena) proporcionalna je vjerojatnosti susreta jedinki predatora i plijena, tj. umnožak njihovih brojeva. U oba slučaja bilinearni članovi pojavljuju se u sustavu modela na desnoj strani odgovarajućih jednadžbi.
Konačno, linearni pozitivni članovi na desnoj strani Volterrinih jednadžbi, koji odgovaraju rastu populacija pod neograničenim uvjetima, odgovaraju autokatalitičkim članovima kemijske reakcije. Ova sličnost jednadžbi u kemijskim i okolišnim modelima omogućuje nam primjenu istih istraživačkih metoda za matematičko modeliranje populacijske kinetike kao i za sustave kemijskih reakcija.
Klasifikacija vrsta interakcija
U skladu s Volterrinim hipotezama, međudjelovanje dviju vrsta čija brojnost x 1 i x 2 može se opisati jednadžbama:
(9.1)
Ovdje su parametri a ja - konstante vlastite brzine rasta vrste, c ja- konstante samoograničenja brojnosti (intraspecifična konkurencija), b ij- konstante interakcije vrste, (ja, j= 1,2). Predznaci ovih koeficijenata određuju vrstu interakcije.
U biološkoj literaturi, interakcije se obično klasificiraju prema uključenim mehanizmima. Raznolikost je ovdje golema: različite trofičke interakcije, kemijske interakcije koje postoje između bakterija i planktonskih algi, interakcije gljiva s drugim organizmima, sukcesija biljni organizmi vezano osobito za natjecanje za sunčeva svjetlost i s evolucijom tla itd. Ova se klasifikacija čini širokom.
E . Odum, uzimajući u obzir modele koje je predložio V. Volterra, predložio je klasifikaciju ne prema mehanizmima, već prema rezultatima. Prema ovoj klasifikaciji, odnose treba ocijeniti kao pozitivne, negativne ili neutralne ovisno o tome povećava li se brojnost jedne vrste, smanjuje ili ostaje nepromijenjena u prisutnosti druge vrste. Zatim se glavne vrste interakcija mogu prikazati u obliku tablice.
VRSTE INTERAKCIJE VRSTA
|
SIMBIOZA |
b 12 ,b 21 >0 |
||
|
KOMENZALIZAM |
b 12 ,>0, b 21 =0 |
||
|
PREDATOR-ŽRTVA |
b 12 ,>0, b 21 <0 |
||
|
AMENSALIZAM |
b 12 ,=0, b 21 <0 |
||
|
NATJECANJE |
b 12 , b 21 <0 |
||
|
NEUTRALIZAM |
b 12 , b 21 =0 |
Zadnji stupac prikazuje predznake koeficijenata interakcije iz sustava (9.1)
Pogledajmo glavne vrste interakcija
JEDNADŽBE NATJECANJA:
Kao što smo vidjeli u predavanju 6, jednadžbe natjecanja su:
 (9.2)
(9.2)
Rješenja stacionarnog sustava:
(1).
![]()
Ishodište koordinata, za bilo koji parametar sustava, je nestabilan čvor.
(2).
![]() (9.3)
(9.3)
C stacionarno stanje (9.3) je sedlo na a 1 >b 12 /S 2 i
stabilan čvor na a 1 12 /s 2 . Ovo stanje znači da vrsta izumire ako je njezina vlastita stopa rasta manja od određene kritične vrijednosti.
(3).
![]() (9.4)
(9.4)
C stacionarno rješenje (9.4)¾ sedlo na a 2 >b 21 /c 1 i stabilan čvor na a 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Stacionarno stanje (9.5) karakterizira koegzistenciju dviju konkurentskih vrsta i predstavlja stabilan čvor ako je zadovoljen odnos:
![]()
Ovo implicira nejednakost:
b 12
b 21
što nam omogućuje da formuliramo uvjet koegzistencije vrsta:
Umnožak koeficijenata međupopulacijske interakcije manji je od umnoška koeficijenata međupopulacijske interakcije.
Doista, neka su prirodne stope rasta dviju vrsta koje se razmatrajua 1 , a 2 isti su. Tada će nužni uvjet za stabilnost biti
c 2 > b 12 ,c 1 > b 21 .
Ove nejednakosti pokazuju da povećanje veličine jednog konkurenta više potiskuje vlastiti rast nego rast drugog konkurenta. Ako je broj obiju vrsta ograničen, djelomično ili potpuno, različitim resursima, gornje nejednakosti su važeće. Ako obje vrste imaju potpuno iste potrebe, tada će jedna od njih biti održivija i istisnuti će svog konkurenta.
Ponašanje faznih putanja sustava daje jasnu ideju o mogućim ishodima natjecanja. Izjednačimo desne strane jednadžbi sustava (9.2) s nulom:
x 1 (a 1 –c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (a 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
U tom slučaju dobivamo jednadžbe za glavne izokline sustava
x 2 = – b 21 x 1 / c 2 +a 2 /c 2, x 2 = 0
– jednadžbe izoklina vertikalnih tangenti.
x 2 = – c 1 x 1 / b 12 + a 1 /b 12 , x 1 = 0
– jednadžbe izoklina vertikalnih tangenti. Točke parnog presjeka izoklina vertikalnih i horizontalnih tangentnih sustava predstavljaju stacionarna rješenja sustava jednadžbi (9.2.), a njihove koordinate ![]() su stacionarni brojevi konkurentskih vrsta.
su stacionarni brojevi konkurentskih vrsta.
Mogući položaj glavnih izoklina u sustavu (9.2) prikazan je na slici 9.1. Riža. 9.1Aodgovara opstanku vrstex 1, sl. 9.1 b– opstanak vrstex 2, sl. 9.1 V– suživot vrsta kada je zadovoljen uvjet (9.6). Slika 9.1Gdemonstrira sustav okidača. Ovdje ishod natjecanja ovisi o početnim uvjetima. Stacionarno stanje različito od nule (9.5) za obje vrste je nestabilno. Ovo je sedlo kroz koje prolazi separatrix, odvajajući područja preživljavanja svake vrste.

Riža. 9.1.Položaj glavnih izoklina na faznom portretu Volterra sustava konkurencije dva tipa (9.2) s različitim omjerima parametara. Objašnjenja u tekstu.
Kako bi se proučavalo natjecanje vrsta, pokusi su provedeni na velikom broju organizama. Obično se odabiru dvije blisko povezane vrste koje se uzgajaju zajedno i odvojeno pod strogo kontroliranim uvjetima. U određenim vremenskim razmacima provodi se potpuni ili selektivni popis stanovništva. Podaci iz nekoliko ponovljenih eksperimenata se bilježe i analiziraju. Istraživanja su provedena na protozoama (osobito cilijatima), mnogim vrstama kornjaša iz roda Tribolium, drosophila i slatkovodnim rakovima (daphnia). Mnogi eksperimenti su provedeni na mikrobnim populacijama (vidi predavanje 11). Pokusi su također provedeni u prirodi, uključujući planarije (Reynolds), dvije vrste mrava (Pontin), itd. Na sl. 9.2. prikazuje krivulje rasta dijatomeja koje koriste isti resurs (zauzimaju istu ekološku nišu). Kada se uzgaja u monokulturi Asterionella Formosa postiže konstantnu razinu gustoće i održava koncentraciju resursa (silikata) na konstantno niskoj razini. B. Kada se uzgaja u monokulturi Synedrauina ponaša se na sličan način i održava koncentraciju silikata na još nižoj razini. B. Tijekom kokultivacije (u duplikatu) Synedrauina istiskuje Asterionella formosa. Navodno Synedra

Riža. 9.2.Natjecanje u dijatomejama. A - kada se uzgaja u monokulturi Asterionella Formosa postiže konstantnu razinu gustoće i održava koncentraciju resursa (silikata) na konstantno niskoj razini. b - kada se uzgaja u monokulturi Synedrauina ponaša se na sličan način i održava koncentraciju silikata na još nižoj razini. V - uz suuzgoj (u duplikatu) Synedruina istiskuje Asterionella formosa. Navodno Synedra pobjeđuje u konkurenciji zbog svoje sposobnosti potpunijeg iskorištavanja supstrata (vidi i predavanje 11).
Eksperimenti proučavanja konkurencije G. Gausea nadaleko su poznati, pokazujući opstanak jedne od konkurentskih vrsta i omogućujući mu da formulira "zakon konkurentske isključenosti". Zakon kaže da u jednoj ekološkoj niši može postojati samo jedna vrsta. Na sl. 9.3. Rezultati Gauseovih pokusa prikazani su za dvije vrste Parametiuma, koje zauzimaju istu ekološku nišu (Slika 9.3 a, b) i vrste koje zauzimaju različite ekološke niše (Slika 9.3 c).
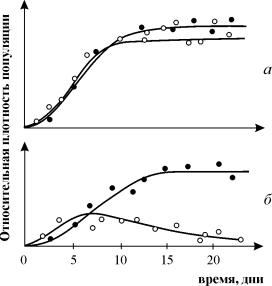
Riža. 9.3. A- Krivulje rasta populacije dviju vrsta Parametij u usjevima jedne vrste. crni krugovi – P Aurelija, bijeli krugovi – P. Caudatum
b- Krivulje rasta P Aurelia i P . Caudatum u mješovitoj kulturi.
Gause, 1934
Model kompeticije (9.2) ima nedostatke, posebice iz njega proizlazi da je suživot dviju vrsta moguć samo ako je njihov broj ograničen različitim čimbenicima, ali model ne pokazuje kolike moraju biti razlike da bi se osigurao dugoročni suživot . Istodobno, poznato je da je za dugotrajan suživot u promjenjivom okruženju nužna razlika koja doseže određenu veličinu. Uvođenje stohastičkih elemenata u model (na primjer, uvođenje funkcije korištenja resursa) omogućuje nam kvantitativno istraživanje ovih pitanja.
PREDATOR+VICTIM sustav
 (9.7)
(9.7)
Ovdje, za razliku od (9.2), znakovi b 12 I b 21 su različite. Kao i u slučaju konkurencije, podrijetlo
![]() (9.8)
(9.8)
je posebna točka tipa nestabilnog čvora. Tri druga moguća stabilna stanja:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Dakle, moguć je preživljavanje samo plijena (9.10), samo predatora (9.9) (ako ima druge izvore hrane) i suživot obiju vrsta (9.11). Već smo raspravljali o posljednjoj opciji u predavanju 5. Mogući tipovi faznih portreta za sustav predator-plijen prikazani su na sl. 9.4.
Izokline horizontalnih tangenti su ravne linije
x 2 = – b 21 x 1 /c 2 + a 1/c 2, x 2 = 0,
a izokline vertikalnih tangenti– ravno
x 2 = – c 1 x 1 /b 12 + a 2 /b 12 , x 1 = 0.
Stacionarne točke leže na sjecištu vertikalnih i horizontalnih tangentnih izoklina.

Od sl. 9.4 vidljivo je sljedeće. Sustav predator-plijen (9.7) može imati stabilan ravnotežni položaj, u kojem o Rumska populacija žrtava potpuno je izumrla ( ) i ostali su samo predatori (točka 2 na sl. 9.4 A). Očigledno, takva situacija može se ostvariti samo ako, osim vrste žrtava o kojoj je riječ, x 1 grabežljivac x 2 – ima dodatne izvore energije. Ta se činjenica u modelu odražava pozitivnim članom na desnoj strani jednadžbe za x2. Posebne točke(1) i (3) (Sl. 9.4 A) su nestabilni. Druga mogućnost – stabilno stacionarno stanje u kojem je populacija grabežljivaca potpuno izumrla i ostaje samo plijen – stabilna točka(3) (Sl. 9.4 6 ). Ovo je posebna točka (1) – također nestabilan čvor.
Konačno, treća mogućnost – održivi suživot populacija predatora i plijena (sl. 9.4 V), čiji su stacionarni brojevi izraženi formulama (9.11).
Kao u slučaju jedne populacije (vidi predavanje 3), za model (9.7) Moguće je razviti stohastički model, ali se ne može eksplicitno riješiti. Stoga ćemo se ograničiti na opća razmatranja. Pretpostavimo, na primjer, da se točka ravnoteže nalazi na određenoj udaljenosti od svake od osi. Zatim za fazne putanje na kojima se vrijednostix 1 , x 2 ostaju dovoljno veliki, deterministički model će biti sasvim zadovoljavajući. Ali ako u nekom trenutku fazne putanje bilo koja varijabla nije jako velika, tada nasumične fluktuacije mogu postati značajne. Oni dovode do toga da se reprezentativna točka pomiče u jednu od osi, što znači izumiranje odgovarajuće vrste.
Stoga se stohastički model pokazuje nestabilnim, jer stohastički "drift" prije ili kasnije dovodi do izumiranja jedne od vrsta. U ovoj vrsti modela, grabežljivac na kraju izumire, bilo slučajno ili zato što je njegova populacija plijena prvo eliminirana. Stohastički model sustava predator-plijen dobro objašnjava Gauseove eksperimente (Gause, 1934), u kojoj treptavice Paramettum candatum poslužio kao žrtva za drugog ciliata Didinium nasatum – predator. Očekivano prema determinističkim jednadžbama (9.7) ravnotežni broj u tim pokusima iznosio je otprilike samo pet jedinki svake vrste, pa ne čudi da su u svakom ponovljenom pokusu ili grabežljivci ili plijen (a nakon njih i grabežljivci) prilično brzo izumrli. Prikazani su rezultati pokusa. na sl. 9.5.

Riža. 9.5. Visina Parametium caudatum i grabežljive trepljarice Dadinium nasutum. Iz : Gause G.F. Borba za egzistenciju. Baltimore, 1934
Dakle, analiza Volterra modela interakcije vrsta pokazuje da, unatoč širokoj raznolikosti tipova ponašanja takvih sustava, u modelu konkurentskih vrsta uopće ne može biti neprigušenih fluktuacija broja. Međutim, takve se oscilacije opažaju u prirodi i u pokusima. Potreba za njihovim teorijskim objašnjenjem bila je jedan od razloga formuliranja opisa modela u općenitijem obliku.
Generalizirani modeli interakcije dva tipa
Predložen je velik broj modela za opisivanje interakcije vrsta, čije su desne strane jednadžbi bile funkcije broja populacija koje su u interakciji. Riješeno je pitanje razvoja općih kriterija za utvrđivanje koje vrste funkcija mogu opisati ponašanje privremene veličine populacije, uključujući stabilne fluktuacije. Najpoznatiji od ovih modela pripadaju Kolmogorovu (1935., revidirani članak - 1972.) i Rosenzweigu (1963.).
 (9.12)
(9.12)
Model uključuje sljedeće pretpostavke:
1) Predatori ne komuniciraju jedni s drugima, tj. stopa reprodukcije predatora k 2 i broj žrtava L istrijebljen u jedinici vremena od strane jednog grabežljivca ne ovisi o g.
2) Povećanje broja plijena u prisutnosti grabežljivaca jednako je povećanju u odsutnosti grabežljivaca umanjenom za broj plijena kojeg su predatori istrijebili. Funkcije k 1 (x), k 2 (x), L(x), kontinuirane su i definirane na pozitivnoj poluosi x, g³ 0.
3) dk 1 /dx< 0. To znači da stopa reprodukcije plijena u odsutnosti predatora monotono opada s povećanjem broja plijena, što odražava ograničenu dostupnost hrane i drugih resursa.
4) dk 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). S povećanjem broja plijena, koeficijent reprodukcije predatora monotono opada s povećanjem broja plijena, krećući se od negativne vrijednosti, (kada nema ništa za jelo) na pozitivno.
5) Broj plijena koje je uništio jedan predator u jedinici vremena L(x)> 0 na N> 0; L(0)=0.
Mogući tipovi faznih portreta sustava (9.12) prikazani su na slici. 9.6:

Riža. 9.6.Fazni portreti Kolmogorova sustava (9.12), koji opisuje međudjelovanje dviju vrsta pri različitim omjerima parametara. Objašnjenja u tekstu.
Stacionarna rješenja (ima ih dva ili tri) imaju sljedeće koordinate:
(1). ` x=0;` y=0.
Ishodište koordinata za bilo koju vrijednost parametra je sedlo (Sl. 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Aodređuje se iz jednadžbe:
k 1 (A)=0.
Stacionarni rješenje (9.13) je sedlo ako B< A (Sl. 9.6 A, b, G), B određena iz jednadžbe
k 2 (B)=0
Točka (9.13) se nalazi u pozitivnom kvadrantu ako B>A . Ovo je stabilan čvor .
Posljednji slučaj, koji odgovara smrti predatora i preživljavanju plijena, prikazan je na sl. 9.6 V.
(3). ` x=B,` y=C.(9.14)
Vrijednost C određena je iz jednadžbi:
Točka (9.14) – fokus (Sl.9.6 A) ili čvor (Sl.9.6 G), čija stabilnost ovisi o predznaku količines
s 2 = – k 1 (B) – k 1 (B)B+P(B)C.
Ako s>0, točka je stabilna akos<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
U stranoj literaturi češće se razmatra sličan model koji su predložili Rosenzweig i MacArthur (1963.):
 (9.15)
(9.15)
Gdje f(x) - stopa promjene broja žrtava x u nedostatku predatora, F( x,y) - intenzitet grabežljivosti, k– koeficijent koji karakterizira učinkovitost prerade biomase plijena u biomasu predatora, e- smrtnost predatora.
Model (9.15) svodi se na poseban slučaj Kolmogorova modela (9.12) pod sljedećim pretpostavkama:
1) broj predatora je ograničen samo brojem plijena,
2) brzina kojom određeni predator jede plijen ovisi samo o gustoći populacije plijena i ne ovisi o gustoći populacije predatora.
Tada jednadžbe (9.15) poprimaju oblik.

Kada se opisuje međudjelovanje stvarnih vrsta, desne strane jednadžbi specificirane su u skladu s idejama o biološkim stvarnostima. Razmotrimo jedan od najpopularnijih modela ove vrste.
Model interakcije dviju vrsta insekata (MacArthur, 1971.)
Model, koji ćemo razmotriti u nastavku, korišten je za rješavanje praktičnog problema suzbijanja štetnih insekata sterilizacijom mužjaka jedne od vrsta. Na temelju bioloških značajki interakcije vrsta napisan je sljedeći model
 (9.16)
(9.16)
Ovdje x,y- biomasa dviju vrsta insekata. Trofičke interakcije vrsta opisane u ovom modelu vrlo su složene. Ovo određuje oblik polinoma na desnim stranama jednadžbi.
Pogledajmo desnu stranu prve jednadžbe. Vrste insekata x jesti ličinke vrste na(član +k 3 y), ali odrasle jedinke vrste na jesti ličinke vrste x ovisno o velikoj brojnosti vrsta x ili na ili obje vrste (članovi –k 4 xy, – y 2). Na malom x smrtnost vrste x veći od prirodnog prirasta (1 –k 1 +k 2 x–x 2 < 0 na malom X). U drugoj jednadžbi član k 5 odražava prirodni rast vrste y; –k 6 y – samoobuzdavanje ove vrste,–k 7 x– jedenje ličinki vrste na vrste insekata x, k 8 xy – povećanje biomase vrste na zbog konzumacije odraslih insekata te vrste na ličinke vrste X.
Na sl. 9.7 prikazan je granični ciklus koji je putanja stabilnog periodičkog rješenja sustava (9.16).

Rješenje pitanja kako osigurati suživot populacije s njezinom biološkom okolinom, naravno, ne može se dobiti bez uvažavanja specifičnosti pojedinog biološkog sustava i analize svih njegovih međusobnih odnosa. U isto vrijeme, proučavanje formalnih matematičkih modela omogućuje nam da odgovorimo na neka opća pitanja. Može se tvrditi da za modele poput (9.12) činjenica kompatibilnosti ili nekompatibilnosti populacija ne ovisi o njihovoj početnoj veličini, već je određena samo prirodom međudjelovanja vrsta. Model pomaže odgovoriti na pitanje: kako utjecati na biocenozu i upravljati njome kako bi se brzo uništile štetne vrste.
Upravljanje se može svesti na kratkoročnu, naglu promjenu vrijednosti stanovništva x I u. Ova metoda odgovara metodama kontrole kao što je jednokratno uništavanje jedne ili obje populacije kemijskim sredstvima. Iz gore formulirane tvrdnje jasno je da će za kompatibilne populacije ova metoda kontrole biti neučinkovita, jer će s vremenom sustav ponovno doći u stacionarni režim.
Drugi način je promijeniti vrstu interakcijskih funkcija između prikaza, na primjer, kada se mijenjaju vrijednosti parametara sustava. Ovoj parametarskoj metodi odgovaraju metode biološke kontrole. Dakle, kada se uvedu sterilizirani mužjaci, stopa prirodnog rasta populacije se smanjuje. Ako u isto vrijeme dobijemo drugu vrstu faznog portreta, onu u kojoj postoji samo stabilno stacionarno stanje s nultim brojem štetnika, kontrola će dovesti do željenog rezultata – uništavanje populacije štetne vrste. Zanimljivo je napomenuti da je ponekad preporučljivo primijeniti utjecaj ne na samog štetnika, već na njegovog partnera. Općenito, nemoguće je reći koja je metoda učinkovitija. To ovisi o dostupnim kontrolama i eksplicitnom obliku funkcija koje opisuju interakciju populacija.
Model A.D. Bazykina
Teorijska analiza modela međudjelovanja vrsta najopsežnije je provedena u knjizi A.D. Bazykina "Biofizika interakcijskih populacija" (M., Nauka, 1985.).
Razmotrimo jedan od modela predator-plijen koji se proučava u ovoj knjizi.
 (9.17)
(9.17)
Sustav (9.17) je generalizacija najjednostavnijeg Volterra modela grabežljivac-plijen (5.17) uzimajući u obzir učinak zasićenosti grabežljivca. Model (5.17) pretpostavlja da intenzitet ispaše plijena raste linearno s povećanjem gustoće plijena, što ne odgovara stvarnosti pri velikim gustoćama plijena. Mogu se odabrati različite funkcije za opisivanje ovisnosti prehrane grabežljivca o gustoći plijena. Najvažnije je da odabrana funkcija raste x asimptotski težio konstantnoj vrijednosti. Model (9.6) koristio je logističku ovisnost. U Bazykinovom modelu hiperbola je odabrana kao takva funkcija x/(1+px). Podsjetimo, to je oblik Monodove formule koja opisuje ovisnost brzine rasta mikroorganizama o koncentraciji supstrata. Ovdje plijen igra ulogu supstrata, a predator igra ulogu mikroorganizama. .
Sustav (9.17) ovisi o sedam parametara. Broj parametara može se smanjiti zamjenom varijabli:
x® (OGLAS)x; g ® (OGLAS)/y;
t® (1/A)t; g (9,18)
a ovisi o četiri parametra.
Za cjelovito kvalitativno istraživanje potrebno je četverodimenzionalni prostor parametara podijeliti na područja s različitim vrstama dinamičkog ponašanja, tj. izgraditi parametarski ili strukturni portret sustava.
Zatim je potrebno konstruirati fazne portrete za svako od područja parametarskog portreta i opisati bifurkacije koje se javljaju kod faznih portreta na granicama različitih područja parametarskog portreta.
Izgradnja cjelovitog parametarskog portreta provodi se u obliku skupa "kriški" (projekcija) niskodimenzionalnog parametarskog portreta s fiksnim vrijednostima nekih parametara.
Parametarski portret sustava (9.18) za fiksni g i mala e prikazan na sl. 9.8. Portret sadrži 10 područja s različitim tipovima ponašanja faznih putanja.
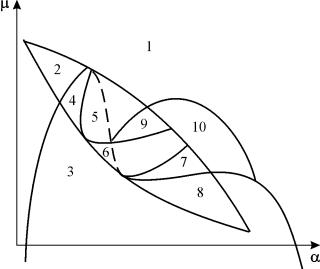
Riža. 9.8.Parametarski portret sustava (9.18) za fiksnig
i mala e
Ponašanje sustava pri različitim omjerima parametara može biti značajno različito (sl. 9.9). Sustav omogućuje:
1) jedna stabilna ravnoteža (regije 1 i 5);
2) jedan stabilni granični ciklus (područja 3 i 8);
3) dvije stabilne ravnoteže (područje 2)
4) stabilan granični ciklus i nestabilna ravnoteža unutar njega (područja 6, 7, 9, 10)
5) stabilni granični ciklus i stabilna ravnoteža izvan njega (područje 4).
U parametričkim područjima 7, 9, 10, područje privlačenja ravnoteže ograničeno je nestabilnim graničnim ciklusom koji leži unutar stabilnog. Najzanimljivija struktura je fazni portret, koji odgovara području 6 u parametarskom portretu. Detaljno je prikazano na sl. 9.10.
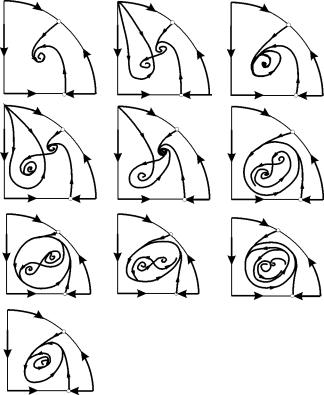
Područje privlačnosti ravnoteže B 2 (zasjenjeno) je "puž" koji se uvija iz nestabilnog fokusa B 1. Ako se zna da je u početnom trenutku vremena sustav bio u blizini B 1, tada je moguće prosuditi hoće li odgovarajuća putanja dosegnuti ravnotežu B 2 ili stabilan granični ciklus koji okružuje tri ravnotežne točke C (sedlo), B 1 i B 2 na temelju probabilističkih razmatranja.
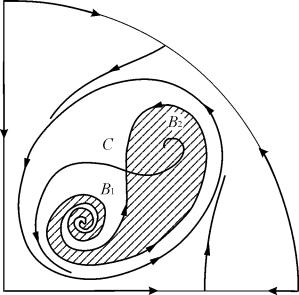
Sl.9.10.Fazni portret sustava 9.18 za parametarsku regiju 6. Privlačna regija B 2 je osjenčana
U parametarskom portretu(9.7) ima ih 22 razne bifurkacijske granice koje tvore 7 razne vrste bifurkacija. Njihova studija nam omogućuje da identificiramo moguće tipove ponašanja sustava kada se njegovi parametri mijenjaju. Na primjer, kada se krećete s područja 1 u područje 3 dolazi do rađanja malog graničnog ciklusa ili laganog rađanja samooscilacija oko jedne ravnoteže U. Slično meko rađanje samooscilacija, ali oko jedne od ravnoteža, naime B 1 , javlja se pri prelasku granica regija 2 i 4. Prilikom napuštanja područja 4 u područje 5 stabilni granični ciklus oko točkeB 1 “puca” na petlji separatrisa i jedina privlačna točka ostaje ravnoteža B 2 itd.
Od posebnog interesa za praksu je, naravno, razvoj kriterija za blizinu sustava granicama bifurkacije. Uistinu, biolozi su dobro svjesni svojstva "puferiranja" ili "fleksibilnosti" prirodnih ekoloških sustava. Ovi pojmovi obično se odnose na sposobnost sustava da apsorbira vanjske utjecaje. Sve dok intenzitet vanjskog utjecaja ne prijeđe određenu kritičnu vrijednost, ponašanje sustava ne doživljava kvalitativne promjene. Na faznoj ravnini to odgovara povratku sustava u stabilno stanje ravnoteže ili u stabilni granični ciklus, čiji se parametri ne razlikuju mnogo od prvobitnih. Kada intenzitet udara prijeđe dopuštenu razinu, sustav se "pokvari" i prelazi u kvalitativno drugačiji način dinamičkog ponašanja, na primjer, jednostavno odumire. Ova pojava odgovara bifurkacijskom prijelazu.
Svaka vrsta prijelaza bifurkacije ima svoje karakteristične značajke, koje omogućuju procjenu opasnosti takvog prijelaza za ekosustav. Evo nekih općih kriterija koji ukazuju na blizinu opasne granice. Kao i u slučaju jedne vrste, ako se pri smanjenju broja jedne od vrsta sustav "zaglavi" u blizini nestabilne sedlaste točke, što se izražava u vrlo sporom vraćanju broja na početnu vrijednost, tada sustav je blizu kritične granice. Pokazatelj opasnosti također je promjena oblika fluktuacija u broju grabežljivaca i plijena. Ako oscilacije koje su bliske harmoničkim postanu relaksacijske, a amplituda oscilacija raste, to može dovesti do gubitka stabilnosti sustava i izumiranja jedne od vrsta.
Daljnje produbljivanje matematičke teorije interakcije među vrstama ide putem detaljiziranja strukture samih populacija i uzimanja u obzir vremenskih i prostornih čimbenika.
Književnost.
Kolmogorov A.N. Kvalitativno proučavanje matematičkih modela populacijske dinamike. // Problemi kibernetike. M., 1972., broj 5.
MacArtur R. Grafička analiza ekoloških sustava // Division of biology report Perinceton University. 1971
A.D. Bazykin “Biofizika interakcijskih populacija.” M., Nauka, 1985.
V. Volterra: “Matematička teorija borbe za opstanak.” M.. Znanost, 1976
Gause G.F. Borba za egzistenciju. Baltimore, 1934.